
Chromosomal Theory of Inheritance
Chromosomal Basis of Inheritance of Class 12
Chromosomal Theory of Inheritance
The salient features of Chromosomal theory (Sutton and Boveri) of inheritance are as follows:
Bridge between one generation and the next is through sperm and ovum. The two must carry all the hereditary characters. Both the sperm and egg contribute equally in the heredity of the offspring. Nucleus contains chromosomes. Therefore, chromosomes must carry the hereditary traits.
Every chromosome or chromosome pair has a definite role in the development of an individual. Loss of a complete or part of the chromosome produces structural and functional deficiency in the organism. Like the hereditary traits the chromosomes retain their number, structure and individuality throughout the life of an organism and from generation to generation. Both chromosomes as well as genes occur in pairs in the somatic or diploid cells. A gamete contains only one chromosomes of a type and only one of the two alleles of a trait. The paired condition of both chromosomes as well as Mendelian factors is restored during fertilization. Genetic homogeneity and heterogeneity, dominance and recessiveness can be suggested by chromosomal type and behaviour. Homologous chromosomes synapse during meiosis and then separate or segregate independently into different cells which establishes the quantitative basis for segregation and independent assortment of hereditary factors.
Chromosomes
Chromosomes (G.K. Chroma - Colour, Soma - body, because of deep staining with certain dyes) are oval, rod or thread like DNA complexes which function as hereditary vehicles.Chromosomes store, replicate and transcribe coded hereditary information (i.e. hereditary vehicles) They were first seen by Hofmeister (1848) as nuclear filaments in pollen mother cells of Tradescantia .Present name was given by Waldeyer (1888)
Prokaryotic Chromosome
In Prokaryotes hereditary material is not associated with proteins. Histones are absent i.e. naked Genetic material contained in a prokaryotic cells is equal to a single chromosome of eukaryotes. It is called nucleoid, genophore, prochromosome or chromonema.
It consists of a single double stranded circular molecule of DNA. In Escherichia coli about 1.2 mm long DNA molecule is accommodated in a space of 1-2 µm. DNA duplex is covalently closed circular structure having several loops each of which is super coiled. Coils are stabilised by RNA and polyamines. Histones are absent but other basic proteins or polyamines are present. Degree of folding is 250-700 times.
A number of genes have been mapped in certain bacteria e.g., 2500 genes in E.coli. The bacterial chromosome attached to mesosome by two closely placed replicating forks.
Mesosome is coiled membranous outgrowth of plasmalemma. Plasmids are small, circular, self replicating extra or additional chromosomal DNA elements, discovered by Hayes and Lederberg (1952).
Plasmids which can get temporarily integrated to genophore and replicate with it are called Episomes. Integration with chromosomal DNA is possible if the plasmid carries some DNA portion similar to chromosomal DNA. The same is called Chromosomal Memory.
Bacterial plasmids carry such factors as fertility, colicinogenic and R factors.
- Fertility factors: Fertility factor (F plasmid) induces conjugation
- colicinogenic factors: Bacteriocidal and Bacteriostatic chemicals are produced by colicinogenic factor (col plasmid)
- R factors: Resistance to certain antibiotics like chloramphenicol, tetracycline, streptomycin and sulphonamide is provided by R-factor (R-plasmid)
These factors are becoming important tools for genetic engineering.
Eukaryotic Chromosomes
In eukaryotes, DNA occurs inside both in nucleus and semi-autonomous organelles (viz. mitochondria, plastids). Eukaryotic chromosomes are DNA-histone complexes. The best time to observe and count the number of chromosomes of a cell is late prophase and metaphase.
- Shape Chromosomes are variable in shape. The shape of chromosome may be V-shaped, L or J-shaped, i-shaped or Rod like.
- Size The size of the normal chromosomes varies from 0.5 µ - 30.0 µm in length and 0.2 µm - 3.0 µm in breadth. Size ofmthe smallest chromosome - 0.25µ e.g. Fungi; Birds etc. Size of the largest chromosome - 30µ e.g. Fritillaria.
Homologous Chromosomes
These are two chromosomes of the same type, size, morphology and arrangement of gene loci.
- Genome It is the complete but single set of chromosomes as found in gametes or gametophytic cells where each chromosome (as well as each gene) is represented singly. Triploid (3n, e.g., Banana), tetraploid (4n, e.g., Rice), hexaploid (6n, e.g., Wheat) ferns and mosses.
- Number The minimum number of chromosomes found in eukaryotes is 2n/2x = 4 (e.g., Haplopappus gracilis, a plant and Ophryortocha puerilis, a polychaete) or n/x = 2(e.g., Mucor hiemalis). A single germ line chromosome is found in roundworm. Parascaris equorum (Ascaris megacephalus var. univalens). Highest number of chromosomes recorded for plants is found in Adder’s Tongue fern (Ophioglossum reticulatum), it is 2n = 1262 chromosomes. Radiolarian protist, Aulocantha, has 2n = 1600 chromosomes.
Sex Chromosomes and Autosomes
These are those chromosomes which singly or in pair determine the sex of the individual in dioecious or unisexual organisms. They are called allosomes or idiochromosomes. A sex chromosome that determines male sex is termed androsome, e.g., Y-chromosome in humans. The normal chromosomes, other than the sex chromosomes if present, of an individual are known as autosomes.
Chemical Composition of Chromosomes
-
DNA
 40% RNA 1.5%
40% RNA 1.5% -
Histone and other basic proteins 50% Acid or NHC proteins 8.5%
-
Lipids traces Ca2+, Mg2+, Fe2+ traces
Ultrastructure of Chromosome
Structurally a chromosome is differentiated into the following parts - pellicle, matrix, chromonemata, primary constriction, secondary constriction, satellite and telomere.
- Pellicle is the outer thin but doubtful covering or sheath of the chromosome.
- Matrix or ground substance of the chromosome is made up of proteins, small quantities of RNA and lipid. It has one or two chromonemata depending upon the state of chromosome.
- Chromonemata (sing.-chromonema) are coiled threads which form the bulk of chromosomes. A chromosome may have one (anaphase) or two (prophase and metaphase) chromonemata. There are three view points about the constitution of chromonema and chromosome.
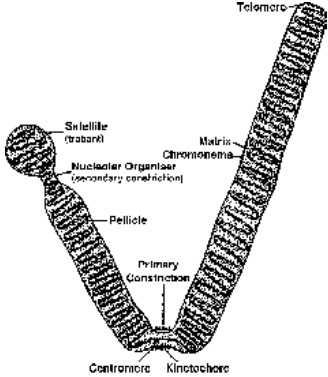
Fig. Structure and parts of chromosome (sat) at anaphase
- Solenoid Model (Finch and Klug, 1976). At intervals, DNA is associated with groups of histone proteins to form nucleosomes having a diameter of 100–125 Å. This beaded string is coiled to form solenoid, like a cylindrical coil of wire used for converting electrical energy into mechanical energy. It has a diameter of 300 Å. The solenoid is coiled into a super-solenoid or chromonema of 2000–4000 Å. The chromonema is folded in the chromosomal matrix to produce a chromatid or chromosome.
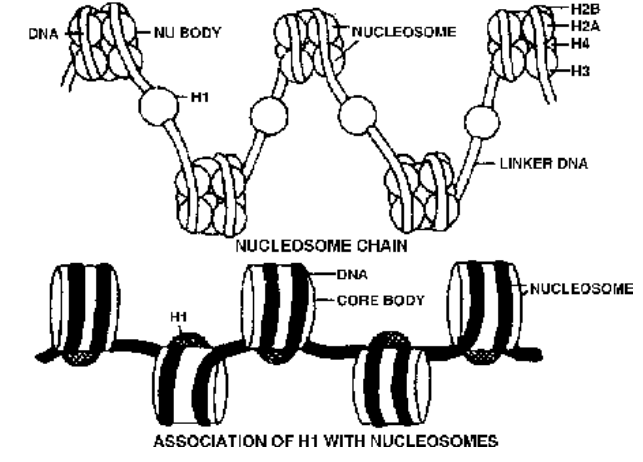
Fig. Nucleosomes and solenoid formation
Histone Proteins
The nucleosome chain is formed by association of DNA with histone proteins. There are five types of histone proteins –
H1, H2A, H2B, H3 and H4. Four of them (H2A, H2B, H3 and H4) occur in pairs to produce histone octomer, nu body or core of nucleosome. They are one of the most conserved proteins, being similar in different species. Their charged ends (+vely) are towards outside. DNA is wrapped over the nu body almost twice to form nucleosome with a diameter of about 100 Å (10 nm). DNA connecting the two adjacent nucleosomes is called interbead or linker DNA. It bears H1. H1 is not conserved but is quite specific for different tissues. Length of linker DNA is also varied.
- Primary constriction is a narrow nonstainable area where the two chromatids are attached in the prophase. The primary constriction is also called centromere. The surface of centromere bears a special trilaminar plate called kinetochore.
- Secondary constrictions are narrow areas other than the primary constriction.
Secondary constrictions are of two types.
- One type of secondary constrictions are produced by breaking and subsequent fusion of chromosome segments.
- The other type of secondary constrictions are metabolically active and function as nucleolar organisers.
In human beings 5 chromosomes (13, 14, 15, 21 and 22) have nucleolar organiser regions. The chromosomes having nucleolar organiser regions also possess satelliltes and are called SAT chromosomes.
- Satellite : The area of a chromosome distal to a nucleolar organiser is called satellite or trabant. It may be small. Commonly a satellite is knob-like in outline. The chromosome bearing a satellite is known as sat chromosome. The word ‘sat’ is not derived from satellite but from poor staining ability of the nucleolar organiser region as its DNA content is low.
- Telomere : The terminal ends of chromosome are named as telomeres. A telomere is a special area of the chromosome having moderately repetitive DNA. It allows chromosome to get attached to the nuclear envelope but not to any other chromosome. Even when a chromosome breaks, the separated segment fuses in the region other than telomere.
Giant Chromosomes Polytene Chromosomes (Kollar, 1882) Polytene chromosomes were first reported by E.G. Balbiani in 1881. They are quite common in salivary glands of insects and are, therefore, popularly called salivary chromosomes.
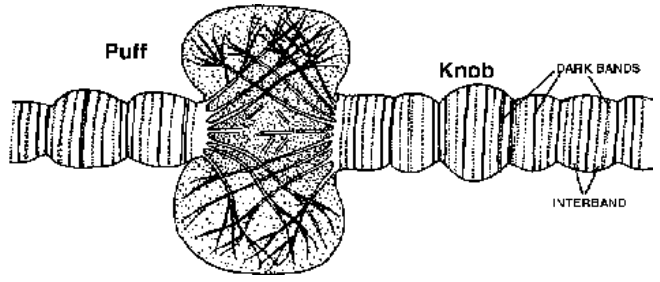
Fig. Enlarged portion of polytene chromosome showing a puff
Lampbrush Chromosomes : The lampbrush chromosomes are highly elongated special kind of synapsed mid-prophase or diplotene chromosome bivalents which have already undergone crossing over.
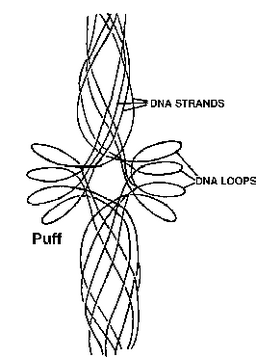
Fig. Interpretation of polytene puff
Functions of Chromosomes
- Chromosomes contain genes. All the hereditary information is located in the genes.
- Chromosomes control the synthesis of structural proteins and thus help in cell division and cell growth.
- Sat chromosomes produce nucleoli for synthesis of ribosomes.
- Some chromosomes called sex chromosomes (e.g., X and Y or X and O) determine the sex of the individual.
Sex Determination
Establishment of male and female individuals or male and female organs of an individual is called sex determination.
It is of three types
- Environmental Determination of Sex : Environment determines sex in some potentially hermaphrodite animals. Marine mollusc Crepidula becomes female when reared alone but produces male in company of an adult (Coe, 1943). Marine worm Bonellia turns female in isolation or attached to substratum. The newly hatched worm develops into male if it happens to get attached to proboscis of female. Such a male enters the female and stays there as a parasite (Baltzer, 1935).
- Nonallosomic Genetic Determination of Sex : In bacteria ‘sex’ determination is controlled by fertility factor located on a plasmid. In several simple organisms, sex is determined by a single gene e.g. Chlamydomonas.
- Allosomic Determination of Sex : Most animals and some plants, sex is determined by sex chromosomes/allosomes. X-body or X-chromosome was discovered by Henking (1891) when he found that in the testes of male bug one chromosome has no homologue.
Stevens (1902) discovered Y-chromosome. Mc Clung (1902) observed that in Squash Bug (Anas), male has 21 chromosomes while female has 22 chromosomes. He stressed that X-chromosome has a role in sex determination.
Wilson and Stevens (1905) put forward the chromosome theory of sex with X and Y chromosomes being named as sex chromosomes.
Correns (1906) put forward the theory of heterogamesis. According to this theory one of the two sexes has different types (heteromorphic) of sex chromosomes. It produces two different types of gametes. The other sex has homomorphic sex chromosomes. It produces one type of gametes or homogametes. Sex of the offspring is determined by the type of heterogamete fusing with the homogamete.
XY Method (XX-XY)
It occurs in mammals and many insects with females having homomorphic XX sex chromosomes and males possessing heteromorphic XY-chromosomes. Y is shorter, may be hooked (as in Drosophila).
XY chromosomes synapse during zygotene. They have two parts, homologous and differential. Homologous regions of the two take part in synapsis. They carry similar genes, called X-Y linked genes, e.g., xeroderma pigmentosum, epidermolysis bullosa. The differential or non-homologous region of Y-chromosome is mostly heterochromatic. The euchromatic part of differential region of this chromosome has holandric genes like TDF (testis determining factor) or SRY *(sex determining region), ZFY (sperm formation), P2V, hypertrichosis (excessive hairiness) of pinna, porcupine skin, keratoderma dissipatum (thickened skin of extremities) and webbed toes. SRY/TDF is the smallest gene (14 base only, Page et al, 1987). Holandric genes are transferred directly from father to son.
Genes present of differential region of X-chromosome also find expression in males (sex-linked traits) whether they are recessive or dominant because males are hemizygous for these genes, e.g., red-green colour blindness, haemophilia. Male is heterogametic (A + Y, A + X, in human beings 22 + Y and 22 + X) while female is homogametic (A + X, in human beings 22 + X). The two types of male gametes or sperms are called androsperms (22 + Y) and gynosperms (22 + X). XY sex determination is found in many bryophytes (females A + X, males A + Y) and angiosperms Melandrium (Lychnis), Coccinia, Salix, Elodea, etc. Sex is decided at the time of fertilization. It is known as syngametic sex determination.
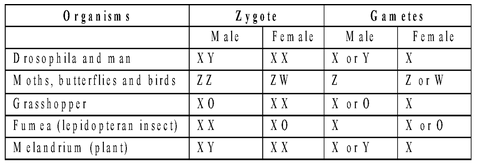
Fig : Sex chromosomal composition of different animals
ZW Method (ZW - ZZ or WZ - WW)
In birds, some reptiles and fishes, the females (hens) have heteromorphic sex chromosomes and are hence heterogametic (A + Z, A + W). W-chromosome determines the female sex. The males have homomorphic sex chromosomes (ZZ) and are hence homogametic (A + Z).
XO Method (XX - XO)
In roundworms and certain insects (true bugs, grasshoppers, cock-roaches), the females have two sex chromosomes, XX while the males possess a single sex chromosome, X. The females are homogametic (A + X). The males are heterogametic producing androsperms (A + Zero) and gynosperms (A + X). Amongst angiosperms, Dioscorea sinuata has XX-XO sex complement.
ZO Method (ZO - ZZ)
In moths and butterflies, the females have odd sex chromosomes (AA + ZO) while males have two homomorphic sex chromosomes (AA + ZZ). The females are heterogametic (A + Z, Z + Zero) and the male homogametic (A + Z).
Haplodiploidy (Honey Bee Method) It is chromosomal sex determination in which one sex is haploid while the other is diploid. Queen Bee picks up all the sperms from a drone during nuptial flight and stores the same in its seminal vesicle. It does not fertilise the eggs which are to develop into drones or males. The phenomenon is called arrhenotoky or arrhenotokous parthenogenesis. Sex determination is progamic in drones and epigamic in females. Drones produce sperms directly without meiosis while queen develops haploid eggs through oogenesis (meiosis). Haplodiploidy also occurs in wasps and ants.
Genic Balance Theory of Sex
It was propounded by Bridges (1926) and is based on his work on Drosophila. According to genic balance theory of Bridges, sex is determined by the balance or ratio between number of X-chromosomes and autosome genomes. Accordingly (XXY + 2A) genotype in Drosophilia produces female phenotype.This is the main proof genic balance theory.
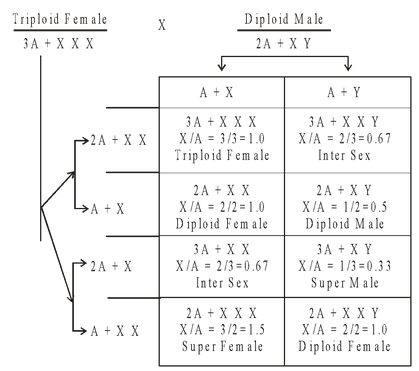
Table : Genic balance concept of sex determination in Drosophila as proposed by Bridges
Chromosome Staining
Chromosomes are stained with special fluorescent dyes that have differential affinity for different parts of the chromosomes. It brings about specific banding pattern. Bands are segments of stand chromosomes that appear lighter, darker or stained as compared to adjacent parts. There are many types of staining techniques which show different banding patterns.
- C-Banding (C-Staining) It stains regions having constitutive heterochromatin (e.g., pericentromeric regions). The chromosomes are first treated with strong alkali. DNA is then denatured with trisodium citrate. It is followed by staining with Giemsa.
- Q-Banding (Q-Staining) Treatment with quinacrine mustard and observation with fluorescent microscope brings out AT rich regions.It was the first fluorescent staining technique developed by Caspersson etal.
- Q-banding This technique stains Y-chromatin in the interphase nucleus of human males.
- G-Banding (Giemsa Staining) Chromosomes incubated in saline solution are stained with Giemsa. It brings out sulphur-rich protein parts. G-Banding is absent in plant chromosomes.
- R-Banding (R-Staining) Chromosomes incubated in buffer at high temperature are treated with Giemsa to produce bands in sulphur deficient regions (hence reverse Giemsa).
Sex Chromatin in Interphase Nuclei
Barr and Bertram (1949) found that interphase nuclei of human females stained with orcein possess small distinct chromatin body called sex chromatin, Barr body or X-chromatin. Number of Barr bodies is one less than the number of X-chromosomes present in an individual, e.g., 1 for normal XX, 2 for XXX.
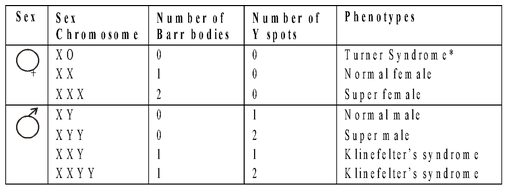
Fig : Sex Chromatin in Interphase Nuclei
Locating Genes on Chromosomes
Morgan and Bridges were the first to experimentally prove that genes are located on the chromosome. The evidence came from two sets of experiments performed independently by them. Morgan searched thousands of Drosophila flies and found a white eyed male fly instead of the normal red colour.
He made three types of crosses.
- Cross 1 : The white eyed male was crossed with red eyed female.
- Cross 2 : Red eyed females of F1 generation were crossed with white eyed males.
- Cross 3 : White eyed females were crossed with red eyed males.

Fig. Cross 1 of Morgan involving red eyed female Drosophila and white eyed male Drosophila.
F1 generation consisted of only red eyed flies. In F2 generation all female flies were red eyed. 50% of the male flies wee red eyed and the remaining 50% white eyed.
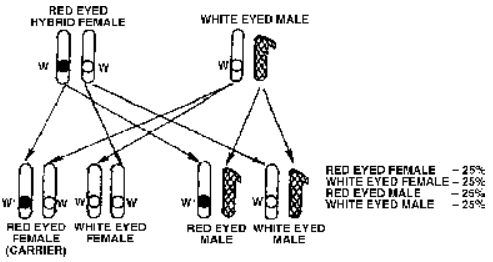
Fig. Cross 2 of Morgan or test cross in Drosophila where red and white eyed traits appear in both males and females in equal proportions

Fig. Cross 3 of Morgan involving white eyed female and red eyed male showing criss-cross inheritance
Taking all the crosses into consideration Morgan came to the conclusion that eye colour gene is linked to sex and is present on the X-chromosome.
Nondisjunction
Bridges (1916) was the first to find out nondisjunction in XX chromosomes in Drosophilia.
- Mitotic nondisjunction : Nonseparation of chromatids in mitotic anaphase is termed as mitotic nondisjunction.
- Primary nondisjunction : The initial nonseparation of synapsed chromosomes that occurs in meiocytes is called primary nondisjunction.
- Secondary nondisjunction : The occurrence of nonseparated chromosomes in the progeny due to previous nondisjunction is called secondary nondisjunction.
