
Microsporogenesis
Share
Microsporogenesis
Reproduction In Flowering Plants of Class 12
Microsporogenesis
This is the process in which each microspore mother cell divides meiotically to form four haploid microspores or pollen grains. These remain arranged in tetrads. The arrangement of pollen grains in a tetrad is affected by cytokinesis during meiosis.
It is of following two types :
- Simultaneous type : In this type, the cytokinesis occurs only at the end of meiosis II. This results in the production of four cells by cellwall formation between the four nuclei. The resultant tetrad shows tetrahedral arrangement and is common in dicotyledons.
- Succesive type : In this type, the cytokinesis occurs twice once at the end of meiosis I, forming two cells and then again at the end of meiosis II to form four cells. The resultant tetrad shows isobilateral arrangement and is found in monocotyledons.
In fact, there are five types of tetrads-
- Tetrahedral
- Isobilateral
- Decussate
- T-shaped
- Linear
In the members of the family Cyperaceae, pollen mother cell divides meiotically to form four nuclei. However, three of these degenerate. The remaining nucleus forms the only microspore. Thus, only one microspore is formed by one pollen mother cell instead of usual four.
In Drosera, Typha, etc., all the four pollen grains do not separate and thus form compound pollen grains. In Mimosaceae,8 to 64 pollen grains remain united together. In Calotropis and some orchids all the pollen grains of an another lobe form a typical structure called pollinium.
Microspore or the pollen grain
Microspore or the pollen grain is the first cell of male gametophyte. It is unicellular and haploid. The shape varies from oval to polyhedral.The wall of the pollengrain is made of two layers. The outer layer is called exine. It is thick and ornamented. At places exine is thin to form germ pores. The exine is made of a substance called sporopollenin . It is biologically the most resistant substance and, hence, pollen grains are the most commonly preserved structures (fossils).
Exine is differentiated into inner endexine and other extexine. Ektexine is formed of 3 layers —
- inner continuous foot layer
- middle discontinuous baculate layer
- outer discontinuous tectum
The inner layer, called intine, is thin and uniform. It is made of pecto-cellulose. At the time of germination, intine comes out of thegerm pores in the formof a pollen tube. In insect pollinated flowers, the exine of the pollen grain is covered with a yellowish viscous and sticky substance called pollenkitt. This is perhaps the protective envelope which also sticks to the body of the insects and thus helps in pollination. The size and form of pollen grains and ornamentation of exine are characteristic of a plant. These are of great taxonomic value and, hence, useful in identifying species. Various characters of pollen grains are studied under a separate branch called palynology.
Microgametogenesis
In this process, a fully developed male gametophyte is formed from a pollen grain or a microspore. Most of the development of malegametophyte is completed inside the anther (in situ) and a part of development occurs on the stigma after pollination. The nucleus of the pollen grain divides mitotically to form a vegetative nucleus (tube nucleus) and a generative nucleus. The later gets surrounded bycytoplasm to become generative cell. At this stage, pollen grain is 2-celled - a large vegetative cell And a small lenticular generative cell.
The pollen grain may be discharged from the anther at this 2-celled stage. However, in some plants generative cell divides further to give rise to two male gametes before the pollen grains are shed. These pollen grains are,thus, 3-celled at the time of shedding.
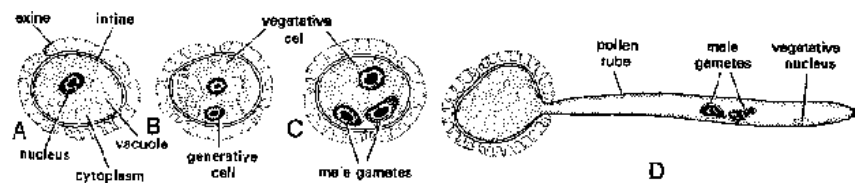
Fig. Different stages of microgametogenesis
Self pollination
This process involves the transfer of pollen grains from the anthers to the stigma of the same flower.
Contrivances or adaptations for self pollination
- Homogamy : In some plants stamens and carpels of a flower mature at the same time. Therefore, pollen grains of a flower can pollinate the stigma of the same flower. Homogamy occurs in Mirabilis, Catheranthus roseus, Gardenia, potato, wheat, rice, etc.
- Cleistogamy : Generally the flowers open after their maturation (anthesis) and expose their anthers and stigma to the environment.Such flowers areknown as chasmogamous flowers. However, some flowers do not open even at maturity and are called cleistogamous flowers. Such cleistogamous flowers are forced to undergo self pollination. Cleistogamy is shown by the underground flowers of Commelina benghalensis (kanchara), Viola mirabilis, etc. Commelina also bears normal flowers on their aerial parts in addition to underground cleistogamous flowers.
Cross pollination

Fig. Agents and types of cross pollination
- Agents and types of cross pollination : Cross pollination involves external agents for the transfer of pollen grains of one flower to the stigma of another flower.
- Biotic agents and Abiotic agents : This agents which include animals of different types such as insects, birds, bats, snails, etc., and abiotic agents like wind and water.
- Entomophily : Pollination by insects is known as entomophily. The common groups of insects bringing about pollination include bees, flies, wasps, drones, moths, butterflies, etc. About 80% of insect pollination is due to bees.
The insect pollinated flowers develop certain adaptations to attract them :
- Large sized flowers
- Bright coloured petals and perianth members
- Scent and odours
- Nectar in special glands
- Edible sap
- Edible pollen
- Special mechanisms
Special mechanism
Salvia often shows Lever or Piston mechanism of pollination. The flower has a bilabiate corolla with two epipetalous stamens. The two anther lobes of a stamen are widely separated by a long curved and lever-like connective. This connective moves freely on the filament. The upper anther lobe is fertile while the lower is sterile. Bees push against the lower anther lobe and swing the connective while entering the corolla tube in search of nectary situated at its base. This brings down the upper another lobe which strikes the back of the insect and dusts it with pollen grains.
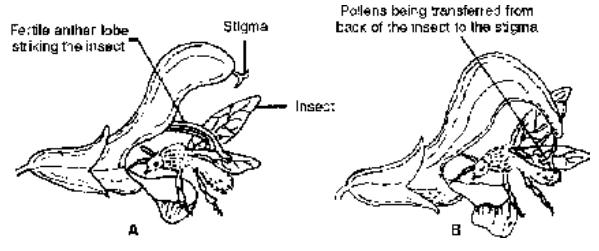
Fig. Pollination in Salvia
Ornithophily : There are only a few bird pollinated flowers. Humming bird and the honey thrushes feed on the nectar of flowers of Bignonia capreolata. Large flowers of Strelitzia raginae are pollinated by sun bird while collecting honey. The plants like Bombax (silk-cotton tree; semal), Erythrina (coral tree),Callistemon (bottle brush), etc., commonly found in the country, are pollinated by crows and mynas.
Cheiropterophily : This is the pollination by bats and commonly occurs in plants growing in tropical regions. Cheiropterophilous flowers are large sized, have long pedicels and produce large amount of nectar. Anthocephalus cadamba (Kadam), Kigelia africana (Sausage tree;), etc., are some of the common bat-pollinated plants.
Malacophily : In these cases pollinating agents are snails and slugs. Land plants like Chrysanthemum leucanthemum and waterplant like Lemna show malacophily.
Anemophily : Wind pollinated or anemophilous flowers are small and inconspicuous. The pollen grains are often dry, small and light in weight. In some cases pollen grains become winged. The style and stigma are generally branched to catch the pollen grains floating in the air. In cereals. ( e.g. , maize) the styles are feathery appearing as tufts of fine, long and silky threads e.g. Gymnosperms, Grasses, Date palm, coconut etc.
Fig. Male Female flower pollination
-
Hydrophily :
The aquatic plants depend upon water for transfer of pollen grains;
e.g.
, in plants belonging to families Najadaceae, Ceratophyllaceae, Potamogetonaceae, Hydrocharitaceae, etc. Pollination may take place in flowers occurring below the water level (Hypohydrophily)
e.g.
Ceratophyllum, Najas, Zostera etc, or in flowers floating on the water sufrace (Epihydrophily)
e.g.
Vallisneria, Hydrilla, Elodea etc.,
Contrivances (adaptations) for cross pollination
- Dicliny or Unisexuality : These flowers are unisexual, i.e. male flowers possess only the stamens and female flowers possess carpels only.
- Dichogamy : This is the condition where stamens and carpels of a bisexual flower mature at different times. Hence, self pollination between them remains ineffective. There are two different conditions
- Protandry : In such flowers anthers mature earlier than the stigma of the flower. Protandry is commonly observed in cotton, lady’s finger, coriander, sunflower, etc.
- Protogyny : In these flowers carpels mature earlier than the anthers of the flower. The common examples include banyan, peepal, ustard apple, Magnolia, Michelia, etc.
- Herkogamy : In some flowers, there is a physical barrier between anther and the style. This prevents self pollination. In members of Cruciferae and Caryophyllaceae, style is long and carries stigma far beyond the stamens. Such a stigma would not receive pollen from the same flower. In Calotropis andsome orchids, pollens occur in special structures called pollinia (Translator). These can be transferred only with the help of insects.
- Heterostyly : In some plants the flowers are of two (dimorphic) or three (trimorphic) forms with anthers and stigmas at different levels. Based on difference in length,the phenomenon is called heterostyly (styles of different lengths) and heteroanthy (stamens of different lengths). Pollination is convenient between stamens and styles of the same length borne by two different flowers. In such flowers there is also a possibility of self-pollination if cross pollination fails. Dimorphic heterostyly is shown by Primula (primrose), Fagopyrum, Linum (linseed), Oxalis (wood-sorrel), etc. Oxalis, Linum and Lythrum show trimorphic heterostyly.
Self sterility
Pollen grains fail to germinate on stigma of the same flower. It is also known as intraspecific incompatibility.
It is of two types :
- Sporophytic incompatibility : It is due to genotype of the sporophytic (stigmatic) tissues.
- Gametophytic incompatibility : It is due to genotype of pollen.
Fertilization
The fusion of nuclei of male and female gametes is called fertilization. This process was discovered by Strasburger (1884). Fertilization occurs in the embryo sac (which lies inside the ovule), the pollen grains received by the stigma have to germinate, produce pollen tube and carry male gametes to the egg cell (siphonogamy).
Germination of pollen grain on stigma and growth of pollen tube
The pollen grain germinates on the stigma by producing a pollen tube which is an extension of intine. Pollen tube comes out through the germ pore. G.B. Amici (1824) discovered the pollen tube in Portulacca oleracea. Generally, only one pollen tube is produced by a pollen grain (monosiphonous). In some plants, for example, members of Cucurbitaceae, many pollen tubes (polysiphonous) are produced. The pollen tube contains a vegetative nucleus or tube nucleus and two male gametes. The male gametes are very small as compared to egg cell. Later, the vegetative cell degenerates. The pollen tube now reaches the ovule after passing through the style.
Entry of pollen tube into the ovule
- Porogamy When the pollen tube enters the ovule through micropyle, it is called porogamy. This method of entry is found in most of the angiosperms.
- Chalazogamy The entry of pollen tube into the ovule from chalazal region is known as chalazogamy e.g. Casuarina.
- Mesogamy When the pollen tube enters the ovule through funicle or integument, it is called mesogamy. e.g. Pistacia,Cucurbita etc.
Entry of pollen tube into embryo sac
The pollen tube enters the embryo sac only from the micropylar end irrespective of its mode of entry into the ovule. The pollen tube either passes between a synergid and the egg cell or enters into one of the synergids through filiform apparatus.
Syngamy and triple fusion
The nuclei of both the male gametes are released in the embryo sac. One of these nuclei fuses with the egg nucleus resulting in the formation of diploid zygote. The process is called syngamy or true fertilization, discovered by Strasburger. The nucleus of the other male gamete enters the central cell and fuses with two polar nuclei or with their fusion product (secondary nucleus). This results in the formation of triploid primary endosperm nucleus. Since, this process involves fusion of three haploid nuclei (two polar nuclei and one male gamete), it is known as triple fusion. The fusion of one male gamete with the egg and fusion of other male gamete with polar nuclei (or their fusion product i.e. secondary nucleus) is called double fertilization. Thus it involves both syngamy and triple fusion. It is unique feature of angiosperms and was discovered by Nawaschin and Guignard in Fritillaria and Lilium.
Significance of double fertilization
The embryo, during its growth and development receives its nourishment from endosperm, which is a product of double fertilization. This process also gives required energy to the polar nuclei for their further development into endosperm, without this process polar nuclei fail to divide further. Since the endosperm nuclei are the resultants of double fertilization, they are characterized by maternal and paternal chromosomes and thus endodperm represents the physiological aggressiveness due to hybrid vigour. The fusion of male and female gametes as well as double fertilization are equally responsible for the production of the viable seeds because absence of any one of these two may cause lethal effect, and the viables seeds are not produced.
Post-fertilization Changes
Following are post-fertilization changes-endosperm formation, embryo formation, seed formation and fruit formation.
- Endosperm (3N) Formation : Endosperm is meant for nourishing the embryo. In Angiosperms, it is triploid because it is formed after vegetative fertilization, where triple fusion takes place. Endosperm may show the effect of genes present in the male gamete.
- Xenia (Focke, 1881) : In plants like Maize, form of endosperm developed depends on male parent i.e., the genes present in male gamete. If the male parent has yellow endosperm and the female parent colourless endosperm then seeds of the product plant will have yellow endosperm.
- Metaxenia (Swingle, 1916) : The effect of pollen on character of the seed coat or pericarp is called as metaxenia.
Types of Endosperms
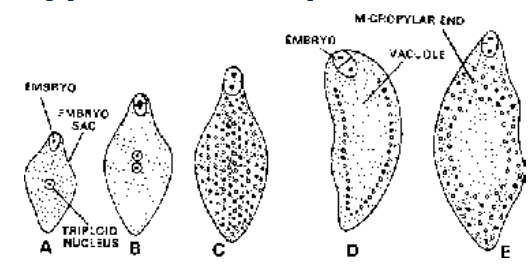
Fig. Nuclear Endosperm
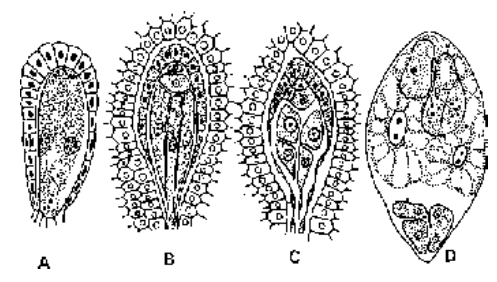
Fig. Cellular Endosperm
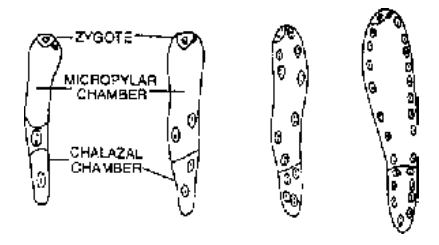
Fig. Helobial Endosperm
Endosperm may persist in the seed. It is called as endospermic seed or albuminous seed e.g. , Castor. It may be consumed by the embryo. In such a case food is generally stored in the cotyledons. Such seeds are called non-endospermic or exalbuminuous., Groundnut, Ruminate or convoluted endosperm occurs in Areca (Betelnut) and Passiflora. Hard endosperm is found in Date, Phytelepas and Areca.
- Nuclear Endosperm : Primary endosperm nucleus divides and redivides to form a large number of free nuclei. A central vacuole appears and a massive peripheral multinucleate cytoplasm is formed. The central vacuole disappears in Capsella bursa-pastoris. Cell wall formation is incomplete in Coconut where there is an outer multicellular solid endosperm and inner free nuclear liquid endosperm in the center.
- Cellular Endosperm : Wall formation occurs after every division of primary endosperm nucleus, so that endosperm is of cellular form from the beginning e.g. , Datura, Petunia.
- Helobial Endosperm : First division of primary endosperm nucleus is followed by cytokinesis to produce two cells; micropylar and chalazal within which free nuclear divisions occurs and cytokinesis takes place at the last step to make them cellular. e.g. , Asphodelus.









