
Secondary Growth In Stelar Region
Anatomy Of Flowering Plants of Class 11
About secondary growth in stelar region
Formation of a Complete Ring of Cambium
In the stelar region, secondary growth occurs by the activity of vascular cambium.
A strip of cambium that originates from the procambium is located between the xylem and phloem of the vascular bundle. This cambium strip is called intrafascicular cambium.
At the onset of secondary growth additional strips of cambium are formed from the interfascicular parenchyma (pith rays) between the vascular bundles. These are known as interfascicular cambium and join with intrafascicular cambium to form a complete ring of cambium.
Structure of Vascular Cambium
The vascular cambium is composed of two types of cells, the fusiform initials and the ray initials.
The fusiform initials are considerably long with tapering ends. Secondary xylem and phloem are derived from these initials.
The ray initials are isodiametric cells and give rise to parenchymatous vascular rays.
Activity of Vascular Cambium
The fusiform initials of the cambium divide periclinally, each forming two daughter cells. One of the daughter cells remains as such so that cambium may remain one cell thick while the other differentiates into xylem or phloem element. The cells which are produced outward form phloem elements, and those inward form xylem elements.
The ray initials of cambium cut plates of parenchymatous cells both outward and inward. These are secondary medullary rays (vascular rays) which extend horizontally from pith to secondary xylem and phloem.
As secondary growth continues, the primary tissues lying outside the secondary vascular tissues, i.e., primary phloem, cortex and epidermis are pushed outside as they are subjected to pressure from inside by the enlarging secondary vascular cylinder and eventually crushed off. The primary xylem persists and can be seen inner to the secondary vascular cylinder.
Secondary Xylem (Wood)
- The secondary xylem forms the bulk of vascular tissue in woody plants. The secondary xylem has two systems of tissues, the axial system and the radial system, which show different orientation in the longitudinal axis of the plant.
- The axial system consists of vertical files of tracheary elements, fibres and wood parenchyma, whereas the radial system consists of rows of parenchymatous cells oriented at right angles to the longitudinal axis of the plant and forms vascular rays.

Fig. Secondary growth in Dicot stem
- Annual rings (Growth rings): In woody plants that grow in temperate regions, the cambium shows marked variation in its activity in different seasons. There is seasonal alternation of periods of activity and quiescence. The wood formed in spring (spring wood) can be distinguished from the one that is formed in autumn (autumn wood) on the basis of differences in shape, structure and distribution of elements in the two regions. The spring wood has much wider and relatively thin walled vessels than the autumn wood. Thus, the part of the secondary xylem (wood) which is added in a single growing period can be easily distinguished from the preceding and succeeding years. The seasonal increments of concentric rings of wood, as seen in transverse section, are said to be annual rings or growth rings. Each ring denotes the growth of one season and from these rings the age of a tree can be calculated. The branch of plant anatomy that deals with determining the age of trees is known as dendrochronology.
- Heartwood and sapwood: In most trees the outer light coloured part of the wood is distinct from the inner dark coloured region. The former is said to be sapwood (alburnum) and the latter heartwood (duramen). The sapwood consists of living cells which are still functional, whereas the heart wood is composed of dead cells which become impregnated with resins, gummy or tannin like substances and consequently are not suited for conduction. As the plant grows older, rings of sap wood bordering heart wood keep on converting into heartwood. This is a gradual process during which the living cells of sapwood lose their protoplast, vessels are blocked by tyloses and there is impregnation of substances like gums, tannin, suberin, etc. Tyloses are tracheal plugs which plug the lumen of vessels.
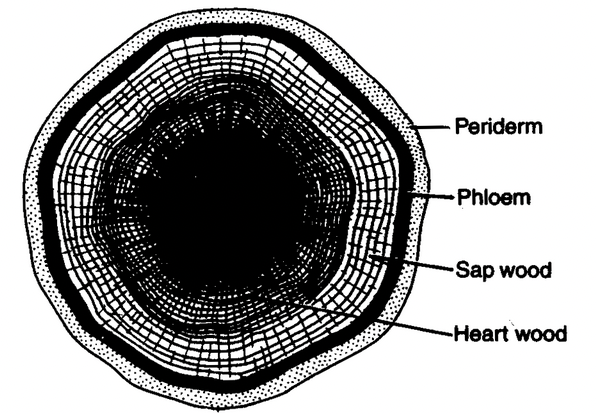
Fig. Heart wood and sap wood in a T.S. of old dicot stem
- Porous and non porous wood: The dicotyledonous wood, which has vessels, is called porous wood, whereas the gymnospermous wood, which lacks vessels, is known as non porous wood. The porous wood is technically referred to as hard wood and non porous as soft wood. This nomenclature has, however, nothing to do with the relative hardness or softness of the wood. Structurally, the hard wood (porous wood) is more complex than the soft wood (non porous wood). In a transverse section of hard wood, vessels appear as small pores. On the basis of distribution of pores within a growth ring, the wood can be classified into ring porous and diffuse porous.
- Ring porous wood: In some angiosperms like Morus and Quercus the vessels formed in the early part of the season are of much larger diameter than the later formed vessels. Thus, rings of wide and narrow vessels occur. This type of wood is said to be ring porous wood.
- Diffuse porous wood: In others, like Acer and Betula, there is no appreciable difference in the size of early and late wood vessels and they are nearly of the same size throughout the season. This type of wood is called diffuse porous wood.
SECONDARY GROWTH IN EXTRASTELAR REGION
A protective tissue usually replaces the epidermis in stems that undergoes pronounced secondary growth. This protective tissue, said to be periderm, is formed by the phellogen or cork cambium. The periderm include the phellogen (cork cambium) and its two derivatives, the phellem (cork) and the phelloderm (secondary cortex).
Phellogen (Cork Cambium)
It is a secondary lateral meristem. In stems, it usually originates superficially in the subepidermal layer or even in the epidermis itself. Histologically, the phellogen is relatively simple and is composed of only one type of cells.
The cells of phellogen appear almost rectangular in a cross section. They are highly vacuolated. The phellogen divides periclinally and produces radially arranged files of cells. The cells toward the outside differentiate as phellem (cork) and towards the inside as phelloderm (secondary cortex).
Phellem (Cork)
The cells of the phellem are dead at maturity and they are characterized by suberin deposits on their walls. They are usually prismatic in shape and are arranged compactly in radial rows. Sometimes, non suberized cells, known as phelloids, also occur in the cork.
The periderm of Quercus suber (cork oak) is the source of commercial cork. The cork is harvested as sheets every 10 years after the tree has attained the age of 20 years. Its light weight, resistance to pressure, thermal insulating qualities, imperviousness to liquids and resistance to acids and other chemicals make it commercially important.
Phelloderm (Secondary Cortex)
The cells of the phelloderm are living cells with non suberized cellulose walls which have simple pits. Chloroplasts may also be present in the cells of phelloderm of some plants. They are often distinguishable from the cells of the primary cortex by their radial alignment with the phellogen and phellem.
Bark
Bark includes all the tissues outside the vascular cambium, i.e., secondary phloem, primary phloem, cortex and periderm. Thus, it is composed of a considerable variety of tissues. If the phellogen forms a complete cylinder around the stem, it gives rise to a ring bark (e.g., Vitis, Clematis) and if it forms arc, it gives rise to a scale bark (e.g. Quercus, Platanus).
Lenticels
In the periderm of most plants, small areas of loosely arranged, thin walled cells are present. These areas are said to be lenticels. They differentiate usually below a stoma. In these regions the activity of the phellogen is more than elsewhere and it forms a mass of loosely arranged, non suberized cells with many intercellular spaces. This tissue is known as complementary or filling tissue. The pressure exerted by the formation of this tissue ruptures the epidermis thus exposing the mass of complementary tissue. Lenticels help in gaseous exchange.
ANOMALOUS SECONDARY GROWTH
The secondary growth that results in the development of a secondary body differing from the common has been termed as anomalous secondary growth.
In most monocotyledons, secondary growth is absent, but in some arborescent monocots like Dracaena, Yucca, Aloe, Agave, Veratrum, Senseviera, Xanthorrhoea, Kingia, etc. an abnormal type of secondary growth takes place. A circular accessory cambium develop in the ground tissue just outside the original vascular bundles. It forms conjunctive tissue and a number of secondary vascular bundles. The secondary bundles may be amphivasal i.e., the xylem surrounds the phloem.
In storage root of sugar beet (Beta vulgaris), sweet potato (Ipomoea batatas), several cambial rings are formed in succession. These cambial rings cut off less secondary xylem and more storage parenchyma in secondary phloem.
Anamolous vascular bundles are those which occur in addition to normal ones like cortical (in cortex, e.g., Nyctanthes) or medullary (in pith, e.g., Boerhaavia).
In some members of family Combretaceae, interxylary or included phloem is found. It is a secondary phloem produced by the abnormal activity of cambium.
INTERNAL STRUCTURE OF LEAF
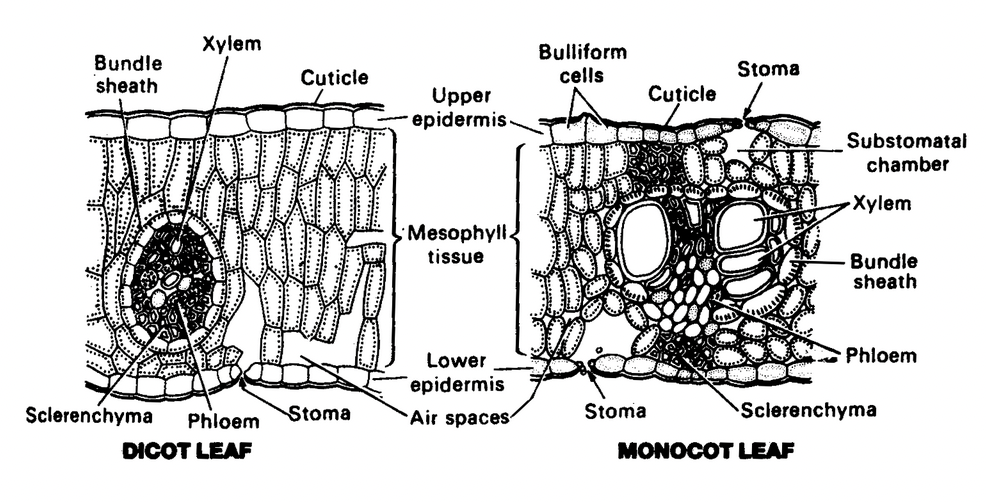
Like the root and stem, the leaf consists of three tissue systems, the dermal system, consisting of the upper and lower epidermis, the ground tissue system, the main photosynthetic tissue which consists of mesophyll, and the vascular system, comprising of veins of various degrees.
Common leaves are bifacial and are further two types, dorsiventral and isobilateral. Unifacial leaves occur in Onion and Garlic.
Epidermis
The upper as well as the lower surface of the leaf is covered by a uniseriate epidermis. However, in some plants (e.g., Nerium, Ficus, etc.) the epidermis is multiseriate.
All epidermal cells of a leaf are alike. The epidermal cells are compactly arranged and their outer walls are usually thickened. The epidermis is covered by a layer of cuticle; the thickness of the cuticle varies considerably and the xerophytic species have a thicker cuticle.
In some xerophytic leaves, especially those of grasses, the epidermal cells situated in longitudinal furrows are large with thin flexible walls. These cells are said to be motor cells or bulliform cells, and they help in rolling of leaves in dry weather.
A characteristic feature of the leaf epidermis is the presence of numerous small openings, called stomata. They occur either on both sides of the leaf (leaf is said to be amphistomatic), confined to the lower surface of the leaf (leaf is known as hypostomatic) or to the upper surface as in floating leaves of aquatic plants (leaf is called epistomatic). A stoma consists of two highly specialized epidermal cells, known as guard cells, enclosing a space. In some plants (e.g., Nerium), stomata present in sunken cavities, called stomatal crypts.
Mesophyll
The bulk of the internal tissue of the leaf, enclosed by the upper and lower epidermis, forms mesophyll. It is composed of thin walled parenchymatous cells containing numerous chloroplasts. The mesophyll is differentiated into palisade and spongy parenchyma in dicot leaves.
The spongy parenchyma consists of irregular and loosely arranged cells, enclosing large intercellular spaces. These air spaces are connected with the substomatal chambers and maintain gaseous exchange with the outside through stomata.
The palisade parenchyma is composed of more or less cylindrical and elongated cells arranged compactly with their long axis perpendicular to the epidermis.
Vascular system
The mid rib (mid vein) in most dicotyledons consists of a single large collateral vascular bundle with an adaxial xylem and abaxial phloem.
The cells surrounding the vascular bundles in the leaf are mostly morphologically distinct from the mesophyll cells. These cells constitute the bundle sheath. In dicotyledons the vascular bundles are surrounded by thin walled parenchymatous cells that extend in the direction parallel to the veins. In monocotyledons, the vascular bundles are completely or partially surrounded by one or two bundle sheaths, each consisting of a single layer of cells.
Anatomical differences between dicot and monocot leaves
|
Character |
Dicot leaf |
Monocot leaf |
|
Type of leaf |
Dorsiventral (bifacial) |
Isobilateral (unifacial) |
|
Stomata |
Mostly present on the lower epidermis |
Equally distributed on both lower and upper epidermis. |
|
Mesophyll |
Differentiated into upper palisade and lower spongy parenchyma. |
Palisade present on both upper and lower surfaces or there is no differentiation of mesophyll into palisade and spongy parenchyma. |
|
Motor cells or Bulliform cells |
Absent |
Present |
|
Supporting tissues |
Parenchymatous |
Sclerenchymatous |
Related Topics
