Pteridophyta
Plant Kingdom of Class 11
The Pteridophytes (Gk. petron = feather, phyton = plant, Haeckel, 1866) are the spore bearing most primitive vascular plants. They are commonly called Vascular Cryptogams.
Oswald Tippo (1942) has placed them in tracheophyta (Sinnot, 1935). The Pteridophytes are an assemblage of flowerless, seedless, spore bearing plants that have successfully invaded the land. They were perhaps the first land plants to evolve during ordovician period (425-450 million years back).
Today the group is represented by about 11,000 living species.
The Salient Features Of Pteridophytes
Occurrence
The Pteridophytes grow under varied habitats, most of them are terrestrial plants and thrive well under damp and shady conditions while some flourish well in open grasslands or even under xeric conditions (e.g., Selaginella lepidophylla). A few pteridophytes grow as aquatic (e.g., Marsilea, Azolla and Salvinia) while still other are epiphytes (e.g., Lycopodium phlegmaria).
The Plant Body (The Adult Sporophyte)
The main independent plant body of pteridophytes is sporophyte (2n).
The sporophytic plant body is cormophyte differentiated into true root, stem and leaves. Some primitive members of the group may lack true root and well developed leaves e.g., order Psilophytales and Psilotales. In these, roots are replaced by rhizoids and stem axis becomes photosynthetic.

Fig.: Different forms of Pteridophytes : A. Psilotum, B. Lycopodium, C. Selaginella, D. Equisetum,
E. Marsilea, F. Adiantum, G. Azolla
The primary root is short lived. It is replaced by adventitious roots which generally arise endogenously.
The stem bears leaves which may be small microphyllous (e.g., Lycopodium, Equisetum) or very large macrophyllous (Pteridium, Pteris and other ferns).
All the vegetative parts possess vascular tissues, organized in definite groups or stele.
The xylem mainly consists of tracheids and xylem parenchyma. The vessels are generally absent but primitive type of vessels have been reported in Selaginella, Equisetum, Pteridium, etc.
The phloem consists of sieve-tube and phloem parenchyma. The companion cells are absent.
Secondary growth does not occur in most of the living pteriodophytes, Isoetes being an exception.
Reproduction
The sporophytic plant reproduces by means of spores that are produced by sporangia.
Leaves bearing the sporangia are called sporophylls.
The sporophylls may be widely scattered on a plant or may be clustered in definite areas and structures called cones or strobili (e.g., Selaginella and Equisetum).
In aquatic Pteridophytes, the sporangia are produced within the specialized structures called the sporocarps (e.g., Marsiliea, Salvinia and Azolla).
According to the mode of development, the sporangia are of two fundamental types, the eusporangium and leptosporangium. The eusporangium is found in most cases and develops from several sporangial initials (e.g., Psilotum, Lycopodium, Selaginella, Equisetum, etc). The leptosporangium is found only in some of the advanced ferns and develops from a single superficial cell (e.g., Marsilea, Pteridium, Pteris, etc.).
Within the sporangia are developed the diploid spore mother cells or sporocytes. These spore mother cells undergo meiosis or reduction division to form spores. If all the spores are of the same size, the plant is said to be homosporous (e.g., most of pteridophytes) and if they are of two different sizes, the plant is called heterosporous (e.g., Selaginella, Isoetes, Marsilea, Salvinia, Azolla). In the heterosporous types, the two different types of spores are produced in separate sporangia. The smaller spores are termed as microspores and are developed in microsporangia, while the larger spores that are generally produced in smaller numbers, are termed as megaspores and are formed in megasporangia.
The Gametophyte
The spores, on germination give rise to the haploid gametophytes or prothalli that are usually small and insignificant structures. The gametophytes are inconspicuous as compared to the sporophytes. One of the most characteristic features of the pteridophytes is that the sporophyte has become the dominant morphological part of the life cycle while the gametophyte has been much reduced.
Sex Organs
The gametophyte or prothallus bears the sex organs, the antheridia and archegonia. Typically, the gametophytes formed from the homospores are monoecious, i.e., both antheridia and archegonia are borne in large numbers on the same gametophyte or prothallus. The gametophytes formed from the heterospores are dioecious, the antheridia and archegonia developing on separate male and female gametophytes.
Antheridia : The antheridia may be embedded either wholly or partially in the tissue of gametophyte or they may project from it. At maturity, each antheridium is a globular structure. It consists of an outer single cell layered sterile wall inside which are found a large number of androcytes. Each androcyte gives rise to a single motile antherozoid.
Archegonia : The archegonia in pteridophytes resemble closely with those of the bryophytes. Each archegonium is a flask shaped structure, consisting of a basal swollen, embedded portion the venter and a short neck. The neck consists of 4 vertical rows of cells, each with 2 to 4 cells. The neck has a single neck canal cell. The venter encloses the egg and ventral canal cell. At maturity, the apical cells separate, the neck canal cell and venter canal cell disintegrate forming a passage for the antherozoids to reach the egg cell.
Fertilization : Fertilization in all cases is accomplished by the agency of water. The fusion of a male gamete and an egg restores the diploid chromosome (2n) number and results in the formation of the zygote.
The Embryo (The New Sporophyte)
The zygote undergoes repeated divisions to form a new sporophyte. The young sporophyte remains attached to the gametophyte by means of a foot and draws nourishment from the prothallus (gametophyte) until it develops its own stem, roots and leaves. The sporophyte is dependent on the gametophyte only during its early stages.
Life Cycle of Pteridophytes
The life cycle of a Pteridophyte comprises of two distinct phases or generations, each of which produces the other. One phase or generation, the sporophyte, is diploid (2n) and the dominant part of the life cycle. It is the part of the life cycle in which vascular tissue is developed. The sporophyte develops sporangia within which are diploid cells called spore mother cells or sporocytes. Each spore mother cell divides by meiotic division. As a result of meiosis, spores are produced which are, therefore, haploid (n).
The gametophytic generation is always small, inconspicuous and bears male and female gametes. The male gamete, i.e., antherozoids are produced in large number within the antheridium. The female gamete, i.e., egg is generally borne singly within the archegonium. Fertilization takes place in the presence of water when an antherozoid fuses with an egg to produce a diploid zygote. The zygote germinates to form the new sporophyte. This generation is called sporophytic generation.
Thus, the life cycle of a pteridophyte consists of an alternate succession of sporophytic and gametophytic generations (heteromorphic or heterologous alternation of generation).
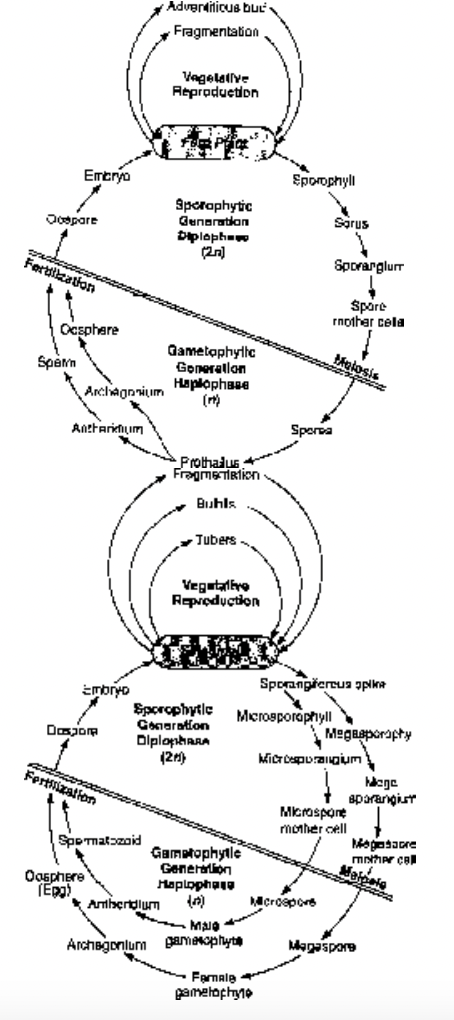
Fig. Life cycle of fern Dryopteris Fig. Life cycle of Selaginella
Stelar System in Pteridophytes
It was given by Van Tieghem and Douliot (1886). Central core of axis is known as stele.
Stele = Xylem + Phloem + Pith (if present) + Pericycle
Different types of steles found in Pteridophytes
1.Protostele This is the most primitive type of stele and all other types have been derived from it.
Here pith is absent. This primitive type of stele is found in Lygodium, Lycopodium, Psilotum and Selaginella sps. etc.
Protostele is of different types:
(i) Haplostele: Here, solid round central core of xylem is surrounded by phloem and pericycle. e.g., Selaginella kraussiana.
(ii) Actinostele: Here, central xylem is star shaped with radiating arms and phloem is present in separate patches e.g., Asteroxylon, Lycopodium serratium, Psilotum, etc.
(iii) Plectostele: Here, xylem is present in form of separate plates and phloem is present between these xylem plates, e.g., Lycopodium clavatum and L.volubile.
(iv) Mixed protostele: Here, xylem is present in the form of discrete units and embedded in phloem, e.g., Lycopodium cernuum.
2. Siphonostele Here, pith is present in the centre of stele, i.e., protostele with pith is called siphonostele or medullated protostele is called siphonostele.
(i) Ectophloic siphonostele: Here, phloem is present only on outer side of xylem, e.g., Osmunda, Equisetum.
(ii) Amphiphloic siphonostele: Here phloem is present on both sides of xylem, e.g., Marsilea, Adiantum.
3. Solenostele Here, the vascular cylinder is broken up by a single leaf gap or siphonostele with single leaf gap is called solenostele.
It is found in some ferns.
4. Dictyostele Here, there are a number of leaf gaps and the vascular cylinder breaks up into a number of smaller units called meristeles and each meristele is protostelic in nature.
It is characteristic of ferns, e.g., Dryopteris, Pteris, etc.
5. Polycyclic stele Here more than one ring of vascular tissue is present. It is found in Pteridium aquilinum, where two concentric rings of vascular tissue are present; outer ring is having a number of meristeles (dictyostele) and inner ring is having only two meristeles. It is also found in Pteris and Marattia.
Heterospory and Seed Habit in Pteridophytes
The occurrence of two kinds of spores in the same plant is called heterospory. Of the two kinds of spores, the smaller ones are called microspores and the larger ones are termed as megaspores and are produced within the microsporangia and megasporangia, respectively.
The production of two kinds of spores in heterosporous plants is definitely related with a differentiation in sex since from a microspore develops a male gametophyte and from a megaspore a female gametophyte. The heterospory, thus, is associated with the sexual differentiation of gametophyte.
The differentiation of spores into microspores and megaspores and their dependence on the parent sporophyte for the nutrition are the certain features in the life cycle of Selaginella that have been considered as the essential prerequisites for the formation of seeds, characteristic of spermatophytes. It is generally agreed that the seed plants arose from the heterosporous vascular plants that instead of discharging the megaspore acquired the habit of retaining it within the megasporangium.
In the seed bearing plants there are two kinds of spores : microspores and megaspores that grow to form male and female gametophytes, respectively. In these plants, the single megaspore is not shed from the megasporangium but is retained within it while still attached to the mother plant. It germinates inside the megasporangium (nucellus) producing the much reduced female gametophyte bearing the archegonia. Later the nucellus and the gametophyte are protected by a covering or integument and the whole structure is known as an ovule. The female gametophyte gets nutrition for its development from the parent plant and thus does not need to produce by its own effort. After fertilization the zygote within the ovule gives rise to an embyro, the rest of the gametophytic tissue gives rise to the nutritive tissue or endosperm and the integument thickens to from a seed coat. This entire structure, i.e., the integumented ovule is known as seed. It is detached from the parent plant and germinates to form a new plant.
Thus we find that for the production of seeds, the following prerequisites are essential :
Production of two types of spores (heterospores).
Megasproangium (nucellus) does not open and the single megaspore within it germinates to form the female gametophyte.
Nucellus becomes invested by a covering or integument which later forms a seed coat.
Within the nucellus is formed a linear tetrad of four haploid megaspores as a result of the reduction division in the functional megaspore mother cell. Out of these four megaspores, the lowermost gives rise to the female gametophyte whereas the rest degenerate.
The male gametes reach the egg by means of a tubular outgrowth of the male gametophyte known as the pollen tube.
Fertilization and formation of embyro take place within the megasporangium.
The embryo undergoes a resting period.
Selaginella exhibits a remarkable approach to the seed habit characteristic of the spermatophytes because of the following features :
The heterospory occurs in almost all the species of Selaginella.
In most species only one functional megaspore mother cell is produced which by reduction division produces four haploid megaspore. In some species, e.g., Selaginella rupestris and S. monospora, only one megaspore is formed in each megasporangium and this single megaspore is not shed but germinates to form the female gametophyte.
The fertilization and development of embryo in both the species take place while the megaspore is enclosed within the megasporangium.
Therefore, it becomes evident that Selaginella is considerably advanced towards the seed habit in a few species but its approach to the true seed is not complete due to the following features :
The megasporangium lacks an integument or covering.
The retention of the megaspore permanently within the megasporangium has not become established.
After the development of the embryo there is a lack of resting period.
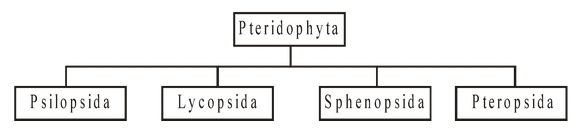
Classification of Peteridophyta
The Pteridophyta is divided into four classes, viz. Psilopsida, Lycopsida, Sphenopsida and Pteropsida, on the basis of organization of plant body including the nature of leaf, vascular system, and location of sporangia.